
")
Post by Omri Jones. Omri is an undergraduate finishing his Bachelor of Science in Psychology, with a minor in Interdisciplinary Neuroscience at Portland State University.

During these past couple months, I have felt lucky to have the opportunity to participate in neuroscience outreach visits to public K-12 schools. During these visits we’ve connected with students, showcased real human brains, and answered many questions that arose.

LEARN MORE: What is outreach like?
Being in a position to consider questions about neuroscience and explore neuroscience topics made me realize how much I don’t know, and how many questions I have about the brain. I found myself more and more curious about a set of brain structures called the basal ganglia that came up in discussions of student questions again and again.
Ever had racing thoughts?
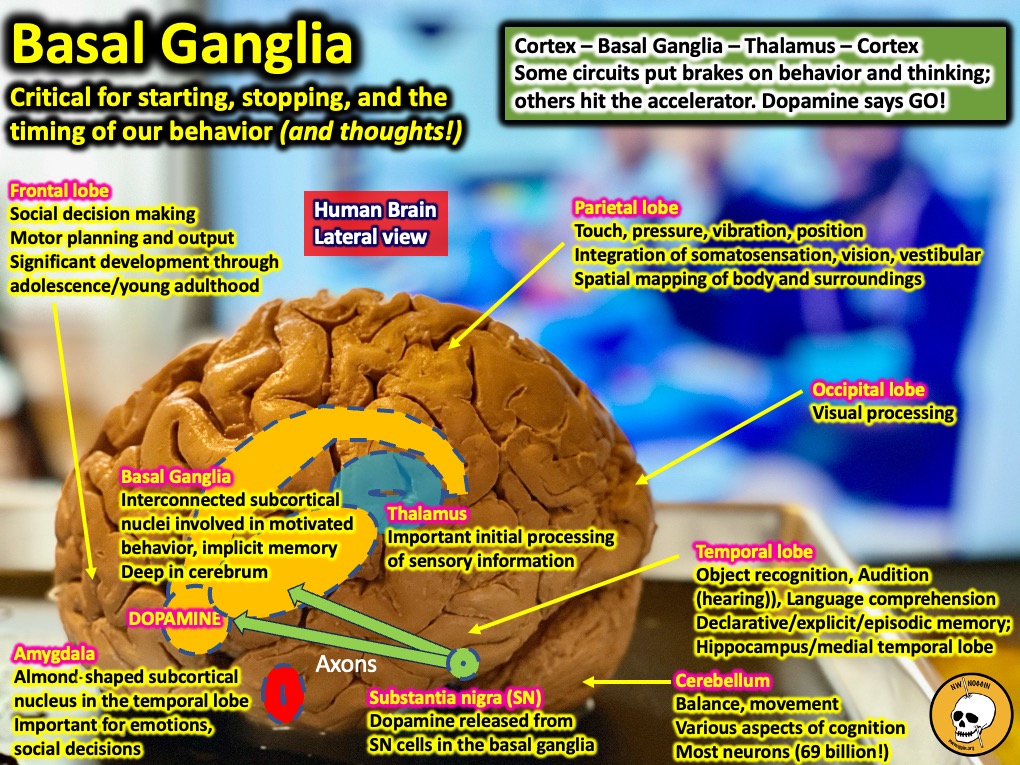
The basal ganglia are a set of grey matter nuclei deep beneath the cortex (subcortical) that are essential for the execution, inhibition, timing and control of movements. In addition to movement, the basal ganglia have been shown to act as a source of control and modulation for a wide variety of cognitive functions, including how fast you’re thinking – and whether or not you can stop.
LEARN MORE: Basal ganglia @ Beaumont!
LEARN MORE: Learning through mistakes!
LEARN MORE: Functional Neuroanatomy of the Basal Ganglia
LEARN MORE: It’s Just Like Riding a Bike…Except It’s Not!
LEARN MORE: A neurosymphony of skills!
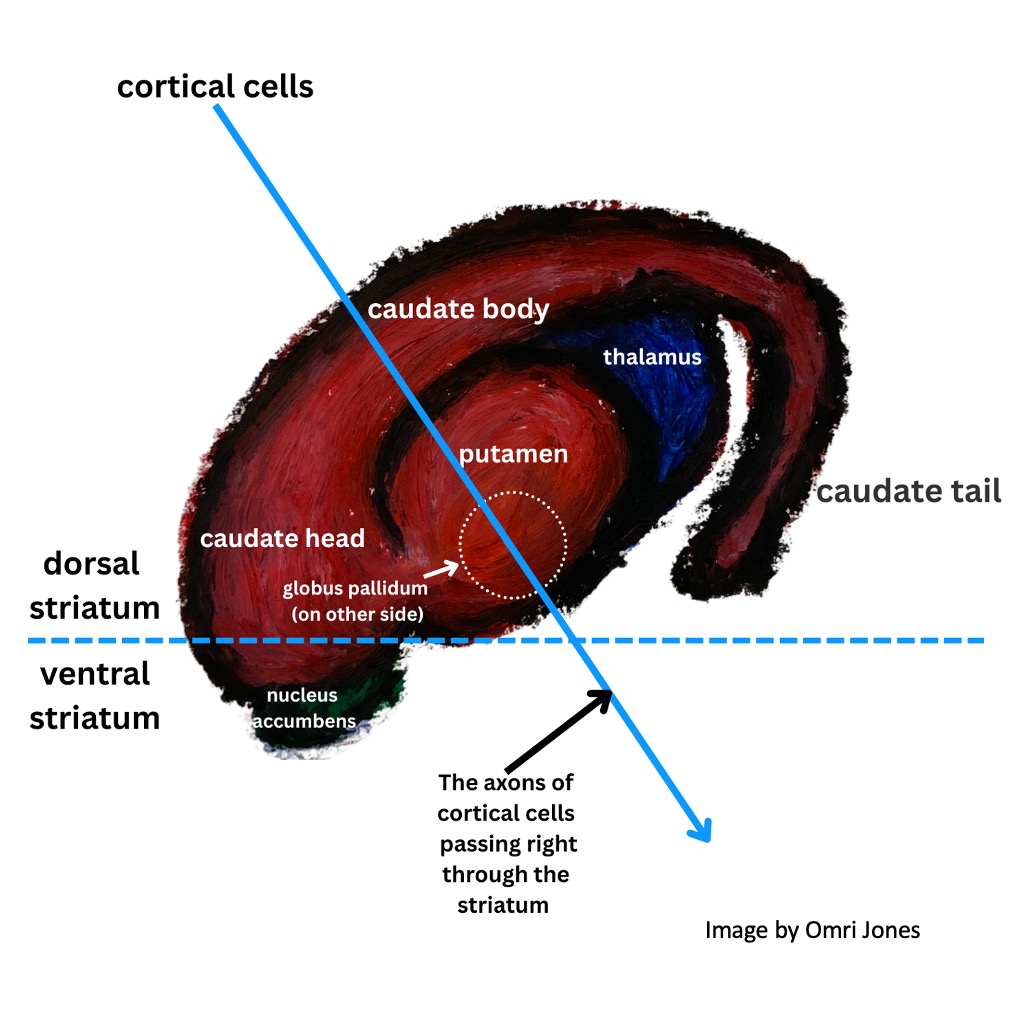
The structures which make up the basal ganglia include two nuclei, the caudate and putamen, which are known collectively as the striatum. Striatum means striated, or striped, because as the tail-like caudate merges with the putamen and another important basal ganglia structure called the nucleus accumbens towards the front of the brain, it gets “striped” by the white matter – the fatty membrane-wrapped (myelinated) axons of cortical motor neurons that pass right through.
LEARN MORE: Neurophysiology and cognitive functions of the striatum
LEARN MORE: The Tail of the Striatum: From Anatomy to Connectivity and Function
LEARN MORE: The Striatum: Where Skills and Habits Meet
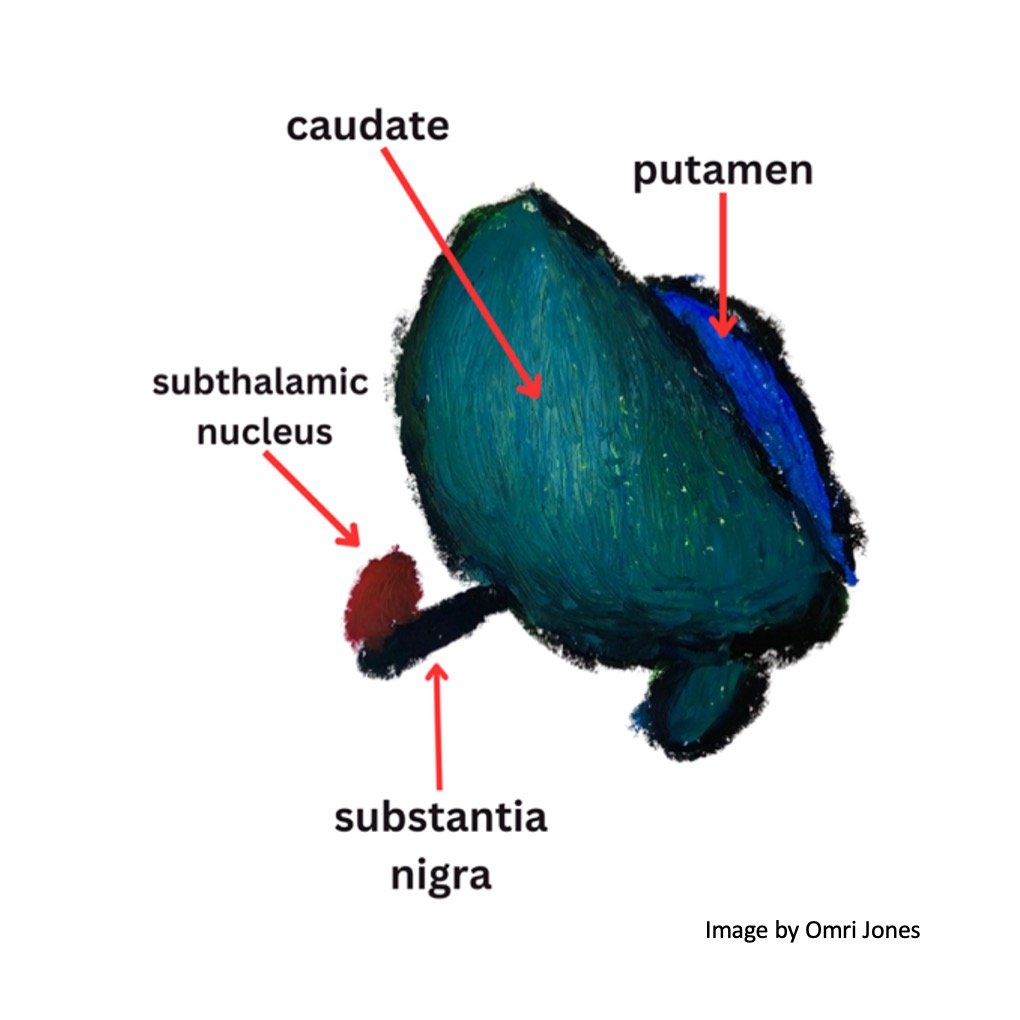
Additional structures in the basal ganglia include the dopamine-releasing substantia nigra (which is located in the brainstem, and implicated in the development of Parkinson’s disease), the subthalamic nuclei, and the globus pallidus (which has both external and internal sections).
Basal ganglia as viewed from the front of the brain.
The caudate has three segments, the head, the body – and a tail!

Basal ganglia viewed from the side of the brain (frontal lobes towards the left).
Something that makes me so curious about the basal ganglia is the elusiveness of their structure.
When looking at a diagram viewed from the side they look completely different than when viewed from the front. The relative sizes of the caudate and putamen to each other also vary based on where a coronal section (a top to bottom slice across the brain’s two hemispheres) is taken from.
Given their complexity and inconsistent appearance in diagrams, I have always felt confused by the way that the basal ganglia appeared differently in different sections of the brain, and I was pleased to have an opportunity to practice identifying these structures on a particular specimen available during outreach.
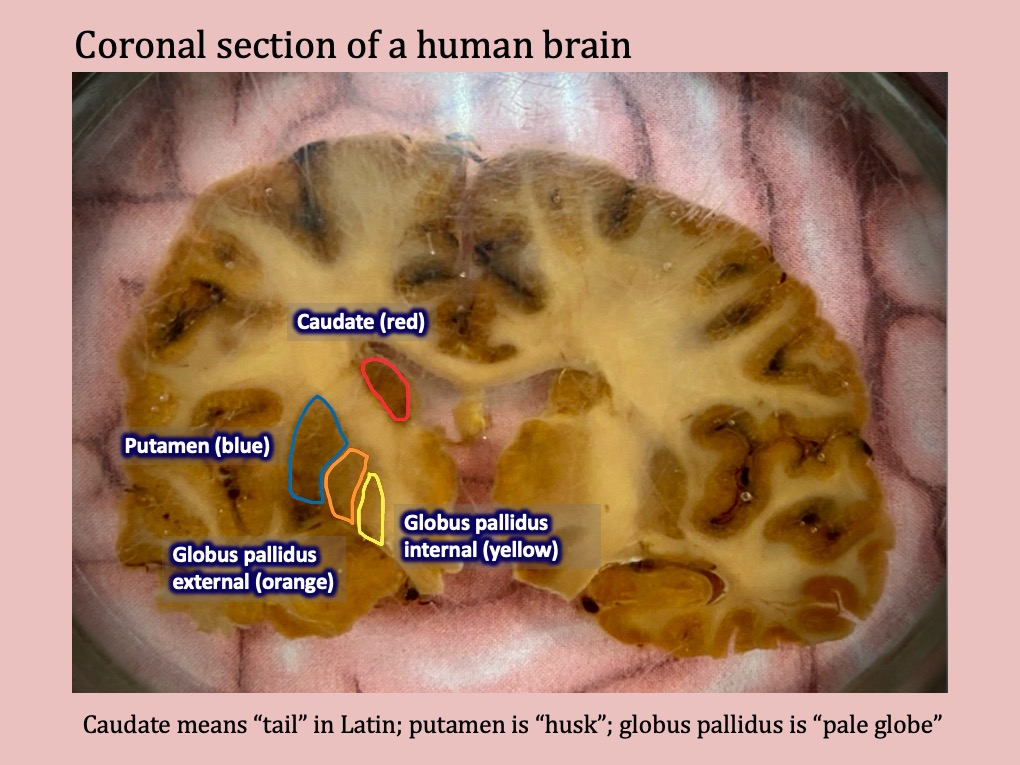
As you can see on this coronal section, the caudate, putamen, and globus pallidus look completely different than they do in both diagrams of the basal ganglia I’ve drawn using pastels above. By looking at this slice it’s hard to imagine that you have the entire set of structures I illustrated in your own brain.
In the same way that being able to identify structures in a real specimen made the content less abstract and much more meaningful for me, students were able to really see what it is that’s doing their thinking, feeling, and living, which I find pretty amazing.

How do the basal ganglia work?
The basal ganglia stop (or inhibit), start and determine the speed of what we do.
Before an action can occur, the cortex signals its intention to the basal ganglia, which then decides whether this action should happen, or if it should be prevented. The signals sent from the cortex must be passed through the basal ganglia to the thalamus, which then relays them back to the same spot in the cortex that they came from. When cortex receives an excitatory signal from the thalamus it allows the intended action to proceed.
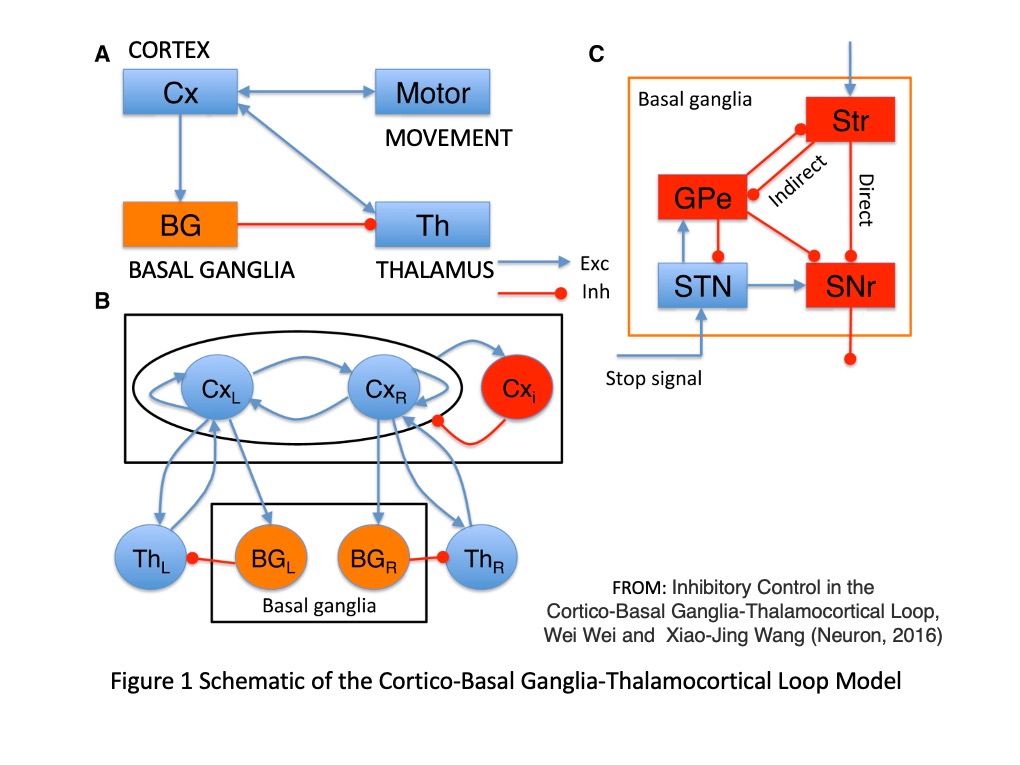
These are complex circuits!
The basal ganglia inhibit the thalamus until they decide to stop, which only then allows for something like a movement (or in some cases even a thought!) to occur. So for action, the nuclei of the basal ganglia that inhibit the thalamus must be turned off by other nuclei in the basal ganglia.
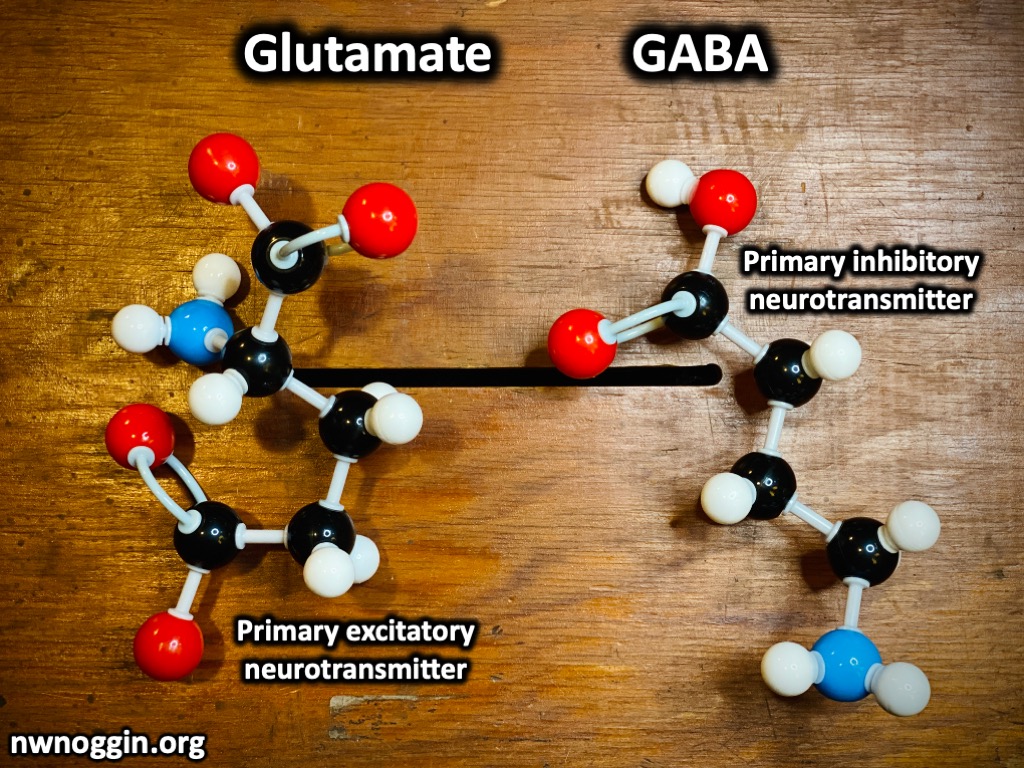
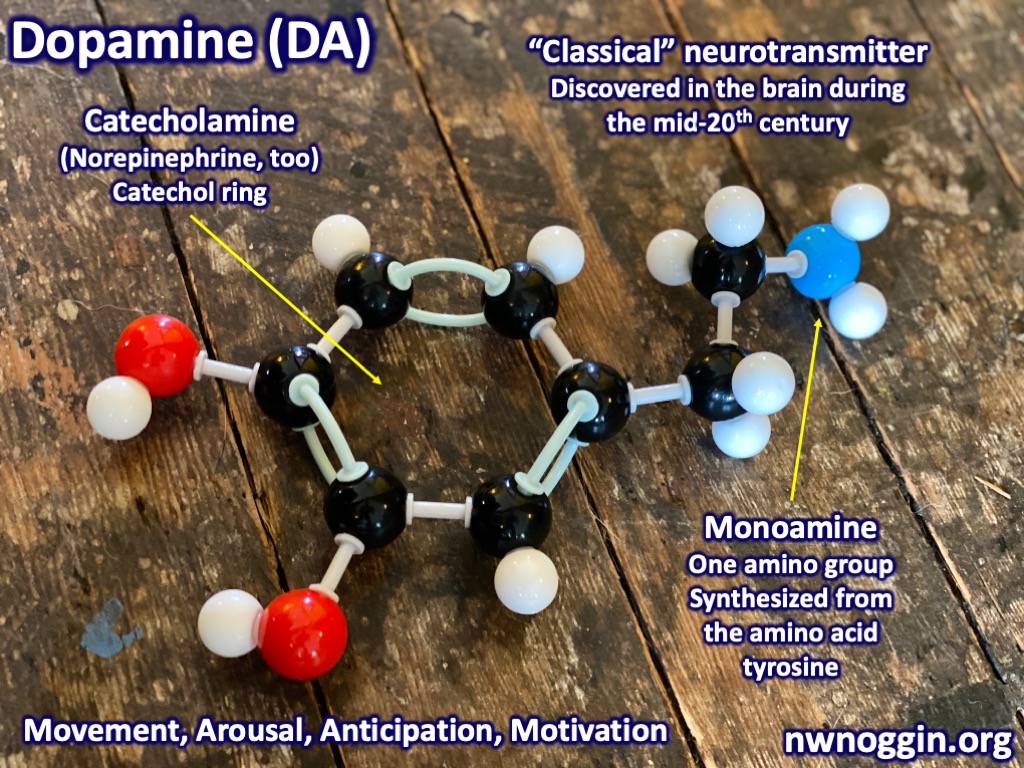
The neurotransmitter used by the basal ganglia to inhibit the thalamus is GABA, and the neurotransmitter involved in the system that releases the thalamus from inhibition is dopamine.
LEARN MORE: Inhibitory Control in the Cortico-Basal Ganglia-Thalamocortical Loop
LEARN MORE: Dynamic control of decision and movement speed in the human basal ganglia
LEARN MORE: The basal ganglia control the detailed kinematics of learned motor skills
LEARN MORE: A Common Function of Basal Ganglia-Cortical Circuits Subserving Speed in Both Motor and Cognitive Domains
LEARN MORE: Untangling the cortico-thalamo-cortical loop
LEARN MORE: The Anatomy of the Basal Ganglia
The substantia nigra has “pars” (parts)!
The substantia nigra (“black substance”) is another critical component of the basal ganglia.
It’s located in the midbrain and consists of two discrete regions: the substantia nigra pars reticulata (the “net-like part”) and the substantia nigra pars compacta (the “compact part”). The pars compacta contains dopaminergic neurons (neurons that release dopamine) and forms the nigrostriatal dopamine pathway, while the reticulata contains GABA-ergic neurons which release inhibitory signals.
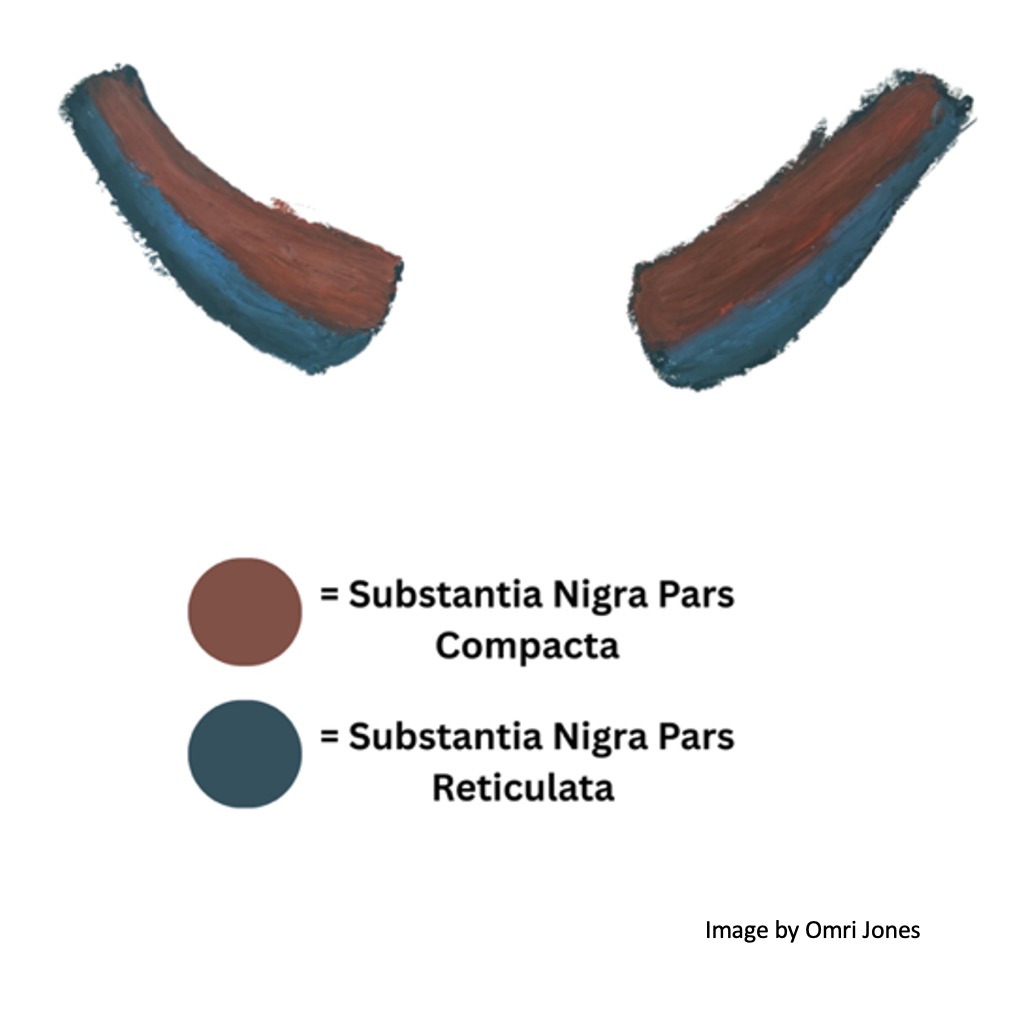
The substantia nigra pars compacta is located above (dorsal to) the pars reticulata and the dopaminergic neuron of the former contain neuromelanin which gives the substantia nigra its dark appearance.
Dopaminergic neurons in the pars compacta send dopamine to the striatum (again, the striped portions of the caudate, putamen and nucleus accumbens) while GABA releasing neurons in the pars reticulata send inhibitory signals to the thalamus and colliculi. As we will soon see, dopamine sent from neurons in the pars compacta releases the thalamus from inhibition and encourages movement, while GABA sent from the pars reticulata inhibits the thalamus preventing movement.
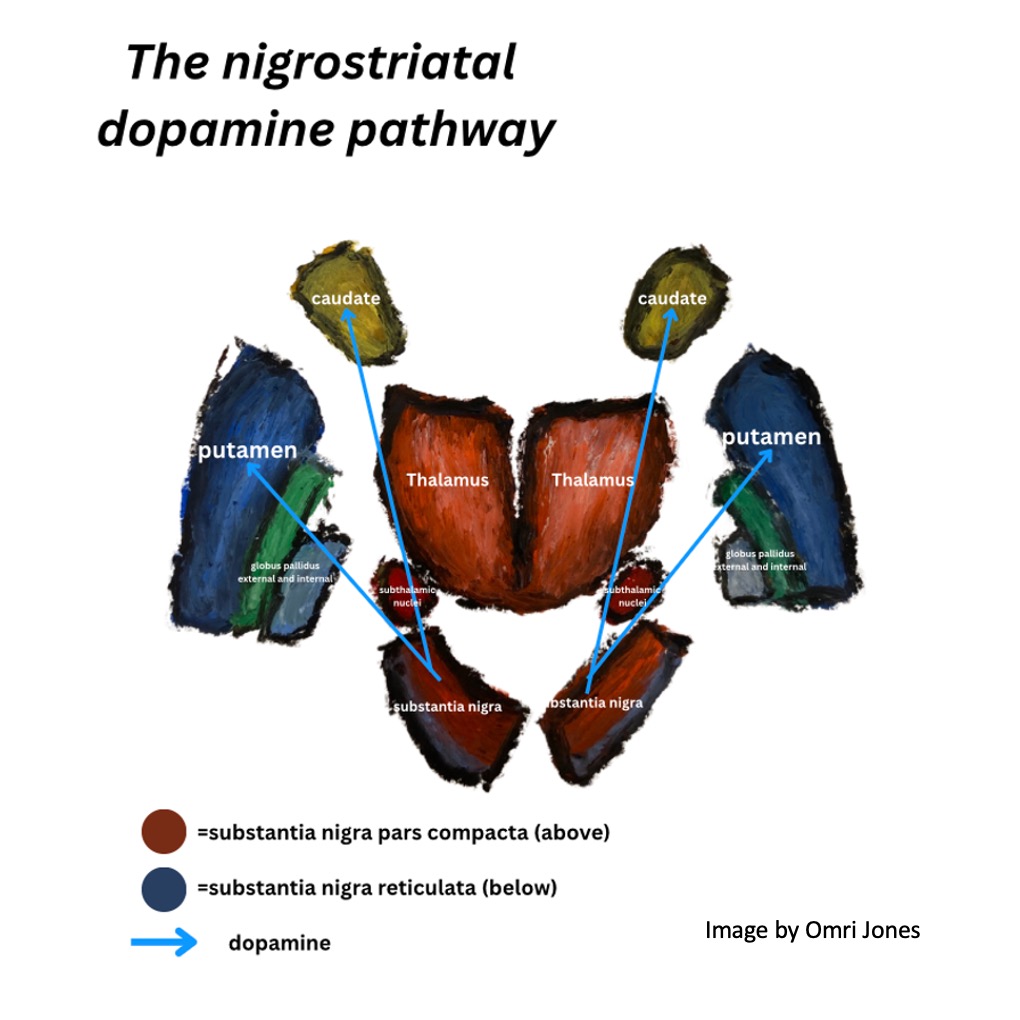
LEARN MORE: Substantia Nigra
LEARN MORE: Neuroanatomy of the Substantia Nigra
Where is dopamine released in the basal ganglia?
The neurons in the striatum are roughly 90% medium spiny neurons that release GABA (an inhibitory neurotransmitter.) Of these neurons there are two populations that differ in their dopamine receptor type, these being D1 neurons and D2 neurons.

IMAGE SOURCE: Dichotomous Anatomical Properties of Adult Striatal Medium Spiny Neurons
D1 neurons in the striatum bind dopamine and become excited, causing them to fire their inhibitory projections onto the internal globus pallidus and substantia nigra pars reticulata. In contrast, D2 neurons in the striatum are inhibited when they bind dopamine, which prevents them from inhibiting their targets which differ from that of the D1 striatal neurons. The D2 striatal neurons project to the external globus pallidus, which inhibits the subthalamic nucleus, which excites the output nuclei needed for inhibition. These different routes give rise to different pathways, termed the direct and indirect pathway.
Direct (go), indirect (stop), HYPER-direct (STOP!)
The direct pathway leads to inhibition of the output nuclei, leading to disinhibition of the thalamus and promotion of a movement. The indirect pathway leads to excitement of output nuclei which increases thalamic inhibition and keeps a movement from happening. And another pathway, called the hyper-direct pathway, cuts off movement quickly, by skipping the striatum altogether.
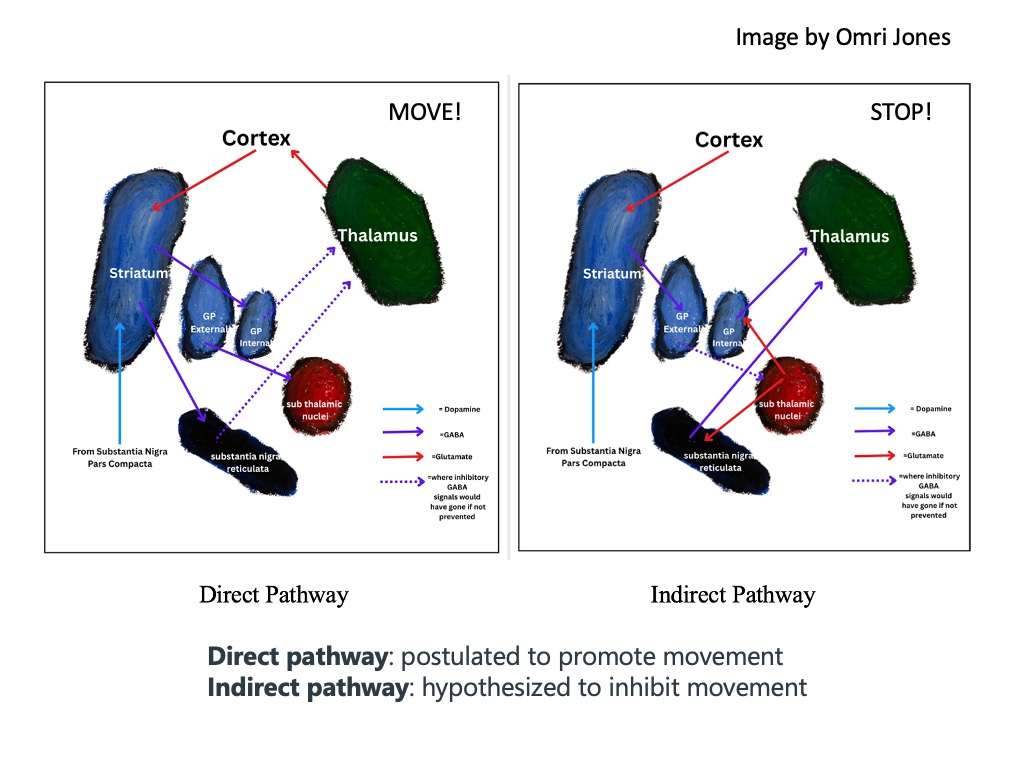
The hyperdirect pathway consists of the cortex exciting the subthalamic nuclei directly by skipping over the striatum. The cortex sends glutamate to excite the subthalamic nuclei, which causes the subthalamic nuclei to release glutamate onto the two principal output nuclei of the basal ganglia (the substantia nigra reticulata and internal globus pallidus.) This excites the substantia nigra reticulata and internal globus pallidus, causing them to very quickly increase their inhibitory output on the thalamus. The hyperdirect pathway is believed to allow for quick stopping of an action that has already been initiated.
In summary, the direct pathway allows an action or thought to occur, while the indirect pathway prevents a thought or action from happening, and the hyper-direct pathway is involved in the QUICK stopping of an action or thought. And dopamine release in the striatum really gets you MOVING!
LEARN MORE: Recent advances in understanding the role of the basal ganglia
Functional divisions of the basal ganglia
The basal ganglia receive input from different regions of the brain in a consistent and functionally segregated way. Information from cortex projects to the striatum, and input from the basal ganglia is then sent to the thalamus, which then relays that information to the exact same spot as where it originated in the cortex.
The nuclei of the basal ganglia can be functionally divided into three distinct divisions based on their consistent patterns of input from the cortex and subcortex.

Associative: The associative division receives projections from regions of the prefrontal cortex and supposedly uses integrated sensory and motor information to influence actions. The associative division is thought to be responsible for goal-directed behavior.
Sensorimotor: The sensorimotor territory receives input from sensory and motor cortices and is thought to modulate motor and sensory output. The sensorimotor division is thought to facilitate habit behavior.
Limbic: This includes projections from the prefrontal cortex, amygdala, hippocampus and other areas involved in emotional responses to the ventral striatum (nucleus accumbens) with modulation by another dopamine releasing nucleus in the brainstem known as the ventral tegmental area (or VTA).
LEARN MORE: Associative and Sensorimotor Cortico-Basal Ganglia Circuit Roles in Effects of Abused Drugs
LEARN MORE: Role of basal ganglia neurocircuitry in the pathology of psychiatric disorders
LEARN MORE: The Brain’s Reward System in Health and Disease
Controlling someone else?
For me, the highlight of these outreach experiences was getting to help facilitate the human-human interface machine.

The Human-to-Human Interface from Backyard Brains is a machine equipped with two sets of electrodes that allow the nerve impulses of one person to be channeled to the muscle cells of another, thus allowing for one person to control the movement of another.


LEARN MORE: Brain Hacking is Electric!
Having tried this experience myself, I felt both humbled due to the fact that this machine only requires a 9-volt battery, and even more curious about the basal ganglia, and what it is to have control over the electrical impulses that you send out to your muscles…and what it is to have control in general.
Insights from disease
Parkinson’s and Huntington’s are two devastating diseases which both involve dysfunction of the basal ganglia. There is a lot of research into these two diseases, and their devastating impacts provide a wealth of information and insight into the way the basal ganglia works.

Parkinson’s disease

Parkinson’s disease (PD) is the result of degeneration of neurons that produce dopamine (dopaminergic neurons) in the substantia nigra pars compacta. Dopamine is required for disinhibition (the removal (“dis”) of inhibition) using the direct pathway. Degradation of these neurons tips the scale towards inhibition of movement and makes it difficult for patients to initiate voluntary movement.
LEARN MORE: What is Parkinson’s disease?
LEARN MORE: Parkinson’s disease explained
Since degeneration occurs in the dopaminergic neurons of the substantia nigra compacta, the dark pigment is lost in patients with PD.

IMAGE SOURCE: Genetic analysis of dopaminergic neuron survival
LEARN MORE: Neuroanatomy of the Basal Ganglia
Huntington’s disease

Huntington’s disease is genetic and caused by the degeneration of medium spiny neurons in the caudate and putamen which is the result of the accumulation of Huntingtin proteins in the striatum. The neurons of the indirect pathway are degraded first, which leads to a reduction in the ability to use the indirect pathway to inhibit, leading to involuntary movement, and cognitive and psychiatric symptoms. The progression of this disease results in the loss of striatal neurons of both pathways, resulting in the complete loss of voluntary movement control, rigidity, and difficulty speaking and swallowing.
Cognitive and psychiatric dysfunction and often noted before motor symptoms. The basal ganglia are thought to be essential for planned and goal directed cognitive behavior, and psychiatric symptoms are thought to be the result of dysregulation in cognitive basal ganglia circuits.
LEARN MORE: Huntington’s Disease
LEARN MORE: Huntington disease: a single-gene degenerative disorder of the striatum
LEARN MORE: The Effect of Huntington’s Disease on the Basal Nuclei: A Review
Control beyond movement
On one of the final outreach visits of the term, a student in one of the classes we visited who had Tourette’s shared with the class some information about her experience, including her family history of Tourette’s, along with some instances where they get worse and what she has found helps her.

Tourette’s Syndrome
The basal ganglia have been shown to be involved in the pathology of Tourette’s syndrome. There is thought to be an increased amount of dopamine being released in the striatum, and dysfunction in the sensorimotor pathway of basal ganglia inhibition. The sensorimotor pathway of the basal ganglia is involved in automatic inhibition of actions and is thought to be central for habit-formation and control of habit behavior. Dopamine is central for habit-formation and driving habit-behaviors.
This increased amount of dopamine released (which favors disinhibition and habit learning), and the dysregulation in regions of the basal ganglia central for habit behavior and the inhibition of automatic actions is thought to lead to tics.
This student described how tics become much more frequent after school, and this made a lot of sense to me, given that the research says that tics become more frequent, and less suppressible with fatigue.
I realized that those with Tourette’s work so much harder to inhibit the impulsive output of the basal ganglia than everyone else, and the combination of energy used for completing the school/workday, and suppressing tics really adds up and leaves less available for suppressing them. Suppressing tics may also lead to more stress, anxiety, and tension, all of which have been shown to worsen tics.
Obsessive Compulsive Disorder
Another DSM diagnosis, obsessive compulsive disorder, has also come up in multiple classes, and considering what we know about the basal ganglia, it is unsurprising to find that the basal ganglia appears to play a role here as well.
The basal ganglia have been shown to provide control and regulation over not just movements, but also thoughts, and other cognitive functions, and significant differences in basal ganglia structure has been demonstrated in a wide variety of conditions.

LEARN MORE: The Basal Ganglia: More than just a switching device
LEARN MORE: Cortico-basal ganglia interactions in movement disorders: Tourette Syndrome
LEARN MORE: Role of basal ganglia neurocircuitry in the pathology of psychiatric disorders
LEARN MORE: Basal ganglia dysfunction in OCD
Motivated and moved
These past couple months of outreach have shown me how much fun it is to share insights into something that’s so relevant to each one of us. I was very pleased to see how many questions students had, and how much they already knew about the brain. The questions also taught me how much I don’t know about the brain, and by interacting with all these neuroscience activities, handling human brains, and talking with students and peers, I started coming up with a lot more questions myself.
My biggest takeaway from these experiences was the extent to which learning goes both ways.

I was inspired by what I learned from all the conversations, the interesting questions, and by identifying structures for students on brain specimens. Additionally, I learned how much fun it is to share neuroscience information with other people, and how curious they are to learn about how our brains work.